
{kind=link}
Demography of SLI elephant seals
Population size
Pinniped population size is not easily estimated, as a portion of the population is at sea at any time. The easiest way to cope with this problem is to estimate the total production of pups and then to use this figure to calculate the actual size of the population using information on the population age structure from life tables. We applied a correction factor of 3.5 to calculate the number of individuals of age one year and older from the number of pups (see below). Therefore, the entire population of Sea Lion Island is estimated to be 1800 seals one year old or older.
Number of breeding females
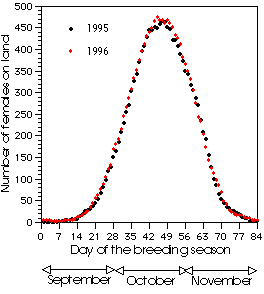
It's quite difficult to obtain directly the total number of breeding females in populations of elephant seals because of the pattern of arrival and departure of females during the three-months breeding season. An exact estimate may be obtained from the marking records of females and their pups: we had 516 in 1995, 518 in 1996, 554 in 1997 and 562 in 1998. These estimates correlate well with direct counts of females (females on land at peak haul out = 90% of the total number of breeding females). A mathematical model of the percentage of females on the breeding beaches in each day of the season was worked out from our daily counts. It is a simple Gaussian model that fits the data much better than other models (e.g., quadratic mode). The fit was very good for all breeding seasons. We used the model to standardize the irregular counts of females carried out prior to our study. These estimates were difficult to compare directly because they had been made in different periods of the breeding season. Since the timing of breeding in our population is almost constant in different years, we used our model to correct the old counts. The population was steady in the 1989-1996 period.
However, current short term stability is not a definitive evidence of stability in the long term, as the small population of Marion Island demonstrates. The other small, localized breeding population of elephant seals for which data is available, at Gough Island, is almost stable. The density of females on Sea Lion Island was quite low (about 110 females per km of the coastline suitable for breeding) compared to densities recorded in other sub-Antarctic populations: this low level of crowding of females probably depends on the small population size and the abundance of sandy beaches with a gentle slope, which are the preferred breeding habitat for elephant seal.
Production and pup mortality
The gross production was almost equal in 1995 and 1996 (517 and 518 pups), while in 1997 we had an increase of 7%. Mortality rate was homogeneous between years, and hence overall estimates can be derived: pre-weaning mortality was 1.6 % and total mortality 2.1%. The mortality was mostly due to still birth.
Mortality increased to 3.4% in 1998. This excess mortality was due to exceptional rough sea during a few days of the season. Pups were frequently washed up and separated by mother, some of them died during high tide, and some other were abandoned. In particular, a small harem of five females, placed in an unusual and marginal site, was completely disbanded, and all females except one lost their pups.
The mortality rates found in our study are lower than those reported for other southern elephant seal populations. In the northern elephant seal the high pup mortality (10-40%) is mainly due to trampling and crushing by the males during their agonistic activities and to starvation through prolonged separation of the pup from the mother because of female aggression or male interference. The same events have been reported for high-density southern elephant seal populations. Visual inspection of dead pups did not offer any indications of crushing by adult individuals; separation of the pup from the mother was always short and in no case definitive. We may conclude that the main sources of mortality observed in crowded populations of elephant seals are of minor importance on Sea Lion Island.
The Sea Lion Island population is a low-density population characterized by medium-sized harems. Low mortality could hence be a result of the low level of agonistic activity between males, the low crowding of females and the low level of female aggression within the harems, as in the Valdes Peninsula.
Sex ratio
On Sea Lion Island, the sex ratio between mature individuals (male of class SAM2 or older) showed a daily variation of Gaussian shape, symmetric for the variation in the number of females on land, and reached a maximum (10-12 females per male) just before the peak haul out of females. The number of females per breeding male (the males which stay on the breeding beaches for prolonged periods actively pursuing access to females) had also a Gaussian daily variation and also reached a maximum (13-15) just before the peak haul out. A stricter criterion for calculating the ratio between breeding individuals is to consider only the alpha males, the males that has control of a harem and, therefore, almost unrestricted access to breeding females (at least in our study population). This may be the best measurement of the actual sex ratio of breeding individuals in a polygynous species with a harem defense mating system. On Sea Lion Island, the overall ratio of breeding females to alpha males reached the maximum (46-47 females per male) just after the peak in the number of hauled out females. The sex ratio between breeding individuals recorded on Sea Lion Island is intermediate between that reported by for the Valdes Peninsula (females/alpha males = 11) and those reported for most sub-Antarctic populations, for example for South Georgia (females/alpha males = 74), for the Kerguelen Islands (102.3), and for Macquarie Island (277).
Male age structure
There was a great variation in the timing of arrival of different age classes on the breeding beaches. Consequently, the age structure of the population on land changed throughout the breeding season. The pattern of males' arrival depended on age; adult males (median day of arrival = day 9 of the breeding season) and older subadults arrived earlier than the younger animals. A peculiar pattern of this variation in age structure was the decrease in the relative number of adult males in the population, a common feature of most elephant seal populations: at peak haulout they represented less than 40% of the breeding males on land. The number of breeding males in each age class increased gradually from subadult class 1 males to adult males (SAM1 = 3.6%, SAM2 = 17.9%, SAM3 = 14.3%, SAM4 = 28.6%, AD = 35.7%). This age structure is typical of a highly competitive mating system in which older males tend to have a higher resource holding potential and tend to keep younger individuals away from breeding areas and females.
Survival
More than half of the breeding females tagged during one season were observed again on Sea Lion Island the next season (67-78%). As females of the genus Mirounga are known to show fidelity to a breeding site, this is likely to be a good estimate of the actual survival rate of females between breeding seasons. We estimated the rate of tags lost by using double-tagged individuals and applying a binomial model of tag loss: tag loss rate are very low. Therefore, the lack of identification of marked females would not significantly change the estimate of female survival between breeding seasons. Nevertheless, the estimate of survival was more accurate for breeding males than for females, as we marked most breeding males with more tags than females, recorded every scar or natural sign on male bodies, and checked each male for signs of lost tags on flippers. We assumed that males not present during the breeding season had died because of the strongly phylopatric nature of the species, because of the lack of evidence that males may skip a breeding season, because of the high quality of our daily resight records and because of the limited chances of breeding outside Sea Lion Islands. About fifty percent of the breeding males survived until the next breeding season.