
{kind=link}
Breeding biology of Sea Lion Island elephant seals
Timing of the breeding season
Reproduction in elephant seals occurs during a fixed three-month period when females come on land to give birth, suckle their pups, mate and return to sea for the post-breeding feeding migration.. The timing of the breeding of pinnipeds is regulated photoperiodically . On Sea Lion Island, the peak presence of females on land was recorded on October 19th or 20th from 1995 to 2017, a remarkable regularity. There is a rough clinal variation of the day of maximum number of females hauled out in various populations of southern elephant seals, and the timing of maximum haul out on Sea Lion Island is in accordance with this cline.
Females begin to come on land during the second week of September. The first birth was recorded on September 11 in 1995 and on September 17 in 1996. The last departure of females was on December 3 in 1995 and on November 27 in 1996: the late departure in 1995 was due to an isolated female with an unusual breeding pattern. On Sea Lion Island, births were recorded over a 60-days period in 1995 and a 58-days period in 1996. These periods are longer than those reported for the Isles Kerguelen (43 days) and Isles Crozet (36 to 51 days), but similar to the times estimated for South Georgia (60 days). On the whole, the length of the birth season seems to be regular both across populations and across seasons in the same population, confirming the rather strict control of timing of breeding in this species.
Female breeding
The timing of female reproduction in land-breeding pinnipeds is usually very regular, and elephant seals are no exception. The typical female comes on land a few days before parturition, gives birth, suckles the pup for about three weeks, mates once or more during a couple of days, and finally returns to sea. Median arrival date was October 5 in each season. Parturition was also very regular, and median date varied from October 10 to October 12. The departure date was more irregular, showing a median between November 1 and November 6. As a consequence of the regular timing of parturition and the predictable interval between parturition and oestrus, the distribution of days of estrus was very similar in the two years, the median date being October 28 or October 29. This regularity probably has a significant effect on male breeding, as the distribution of females in oestrus is very predictable both in space and time.
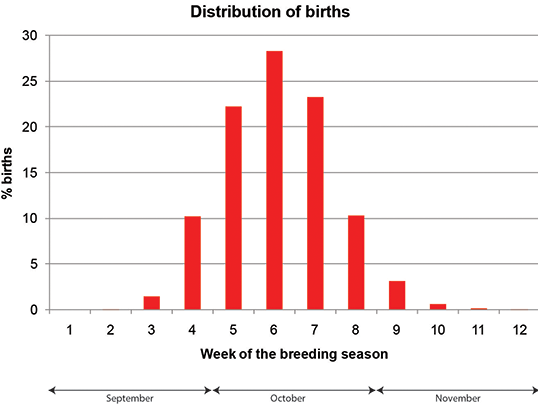
The total time spent on land (arrival to departure) was very similar among females, showing a median of 27 days. The interval between parturition and the beginning of oestrus (first non-protested copulation) was also regular with a median of 20 days. This regularity is expected to promote synchronization in female mating and the predictability of the distribution of fertile periods over the breeding season.
The timing of reproduction in our study area is almost the same as the one reported for the Valdes Peninsula and South Georgia. The main difference between the populations is the day the season starts, while internal timing is almost equal. Female phenotype and timing of reproduction Preliminary evidence suggests that the timing of reproduction partly depends on some aspects of the female phenotype, particularly size. Small females tended to arrive earlier than large females (median arrival day = 29 vs 34), with medium females closer to large ones. Small females also tend to give birth earlier than large females (median parturition day = 34 vs 40). This variation in the timing of breeding of females of different age classes suggests a variation in breeding strategies based on phenotypic differences. Large females are expected to have greater fat reserves and hence to be able to stay on land and suckle their pup for a longer period. Small and medium females spent a lower median number of days on land than large females (27 days vs 29 days). Small and medium females spent a lower median number of days suckling their pups than large females (21 days vs 23 days). Large females seem to be able to stay on land longer and to suckle their pups for more days than smaller females. The absolute value of the difference in timing is small, but suckling is a heavy cost for the mother, as the energy transfer rate between mother and pup is very high. Therefore, even one additional day of suckling may have a significant impact on the female's energy consumption and the weaning weight of the pup.
Sex ratio at weaning
In our study area, the sex ratio was slightly biased toward males (0.87 to 0.94 females per male), although the difference from an equal sex ratio was not significant. A slight bias toward males at birth is common in the majority of elephant seal populations, ranging from 0.82 in South Georgia to 0.89 in the Kerguelen and 0.90 on Heard Island, although an almost balanced sex ratio may sometimes be found. Classical theory of evolution of breeding effort predicts an adaptive variation of offspring sex ratio of different mothers with different phenotype. We compared sex ratio at birth of Sea Lion females classified by size and found small, non significant differences between size classes. This is rough, preliminary evidence against adaptive variation of sex ratio at birth according to capability of maternal investment. Previously published data are contradictory; no adaptive shift in parental investment was reported in M. angustirostris, while for M. leonina there was variation between studies.
